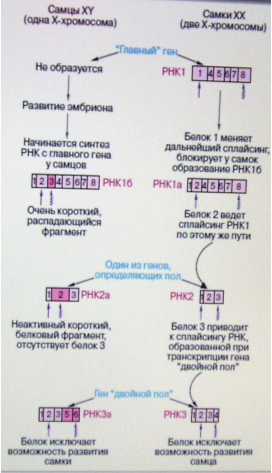
Рис.13. Регулируемый каскад следующих друг за другом альтернативных путей сплайсинга определяет становление пола на самых ранних стадиях развития дрозофилы. Прямая и волнообразная стрелки обозначают соответственно начало и конец трансляции (образование белка). Экзоны, специфичные для самцов, выделены розовым цветом.
У дрозофилы (впрочем, как и у человека) есть две половые хромосомы (X и Y). Две X-хромосомы имеют самки, тогда как самцы - единственную X-хромосому, а также Y-хромосому. Наличие двух X-хромосом у самок достаточно для включения "главного гена", заставляющего включиться "нижележащие" гены, участвующие в определении пола. Главный ген (рис. 13) поначалу начинает работать (транскрибироваться) только у самок с двумя Х- хромосомами. В клетке срабатывают механизмы, подсчитывающие число Х-хромосом. Этот механизм основан на том, что у самок с двумя Х-хромосомами образуется достаточное количество белка, кодируемого генами Х-хромосомы, который активирует промотор главного гена. Транскрипция главного гена начинается с промотора 1 (на рис. 13 не показан). У самцов с одной Х-хромосомой количество белка недостаточно, чтобы запустить главный ген. В результате сплайсинга транскрипта главного гена образуется РНК 1, содержащая экзоны 1, 4, 5, 6, 7 и 8. В этом случае потенциальные экзоны 2 и 3 не входят в состав мРНК. Эти события происходят на самых ранних стадиях развития эмбриона. Немного позднее ген 1 начинает транскрибироваться как у самок, так и у самцов с использованием другого промотора 2, лежащего левее. В этом случае 5'-конец РНК-предшественника будет удлинен, в результате чего экзон 1 сплайсируется иным путем и становится короче в РНК 1а и 1б по сравнению с РНК 1. Белок 1, образующийся при трансляции РНК 1, является регуляторным белком сплайсинга, отсутствующим у самцов. Этот белок направляет сплайсинг у самок с образованием зрелой РНК 1а, содержащей фрагмент экзона 1, к которому пришиваются последующие экзоны: 2-4-5-6-7-8. Белок, определяющий такой характер альтернативного сплайсинга, отсутствует у самцов. В результате в ядрах клеток развивающегося эмбриона, обладающего только одной Х-хромосомой, предшественник РНК созревает с образованием РНК 1б, включающей экзон 3.
Регуляторный белок 1 сплайсинга, образовавшийся у самок на РНК1, позволяет машине сплайсинга "узнавать" в клетках самок экзон 3 как интрон и удалять его с образованием РНК1 и РНК1а. Оказывается, экзон 3 содержит так называемый стоп-кодон, который не кодирует аминокислоты и останавливает рост полипептидной цепи белковой молекулы. В результате синтез белка на РНК 1б если и начнется, то быстро оборвется с образованием очень короткого, распадающегося белкового фрагмента. Таким образом, "самцовый" тип сплайсинга приводит фактически к инактивации гена, поскольку ген не способен кодировать образование функционирующей белковой молекулы. Следовательно, результатом альтернативного сплайсинга может быть не только образование изоформ белков, кодируемых одним геном, но и инактивация гена ("главного гена" у самцов). Белок 2, образующийся у самок, не только постоянно направляет ход сплайсинга с образованием РНК 1а, но и регулирует созревание РНК 2, содержащей экзоны 1-2-3. Эта РНК 2 образуется уже на другом гене, входящем в систему генов определения пола у дрозофилы. У самцов образуется несколько иной вариант РНК 2 - РНК 2а, включающая несколько увеличенный экзон 2. Этот фрагмент экзона у самок узнается как интрон. Опять "самцовая" часть экзона 2 содержит стоп-кодон, останавливающий трансляцию, тогда как у самок образуется активный белок 3, приводящий к образованию РНК 3, включающей экзоны 1-2-3-4. При отсутствии белка 3 предшественник РНК 3 в ядрах клеток самцов "сплайсируется" таким образом, что последовательность, соответствующая экзону 4 самок, выглядит как интрон, а к экзону 3 пришиваются экзоны 5 и 6. На этот раз РНК 3а (экзоны 1-2-3-5-6) у самцов, кодируемая геном "двойной пол", не содержит вредных стоп-кодонов, она кодирует функциональный белок. Мишенями действия регуляторных белков, синтезированных на матричных РНК 3 и 3а, являются гены, дальнейшая работа которых полностью исключает соответственно развитие самца или самки (рис. 13). Отметим, что, несмотря на вариации в характере сплайсинга, всегда по краям интронов должны находиться упомянутые выше канонические нуклеотидные последовательности, используемые при катализе того или иного пути сплайсинга. Мы проследили каскад регулируемых событий альтернативного сплайсинга. У самцов он дважды приводил к образованию дефектной мРНК, не способной кодировать достаточно длинный и функционирующий белок. У самок наблюдается последовательное, поэтапное образование матриц РНК для кодирования белка, регулирующего транскрипцию и/или ход событий альтернативного сплайсинга.
Перейти на страницу:
1 2 3