Оказалось, что экзоны, если их несколько, могут сшиваться (сплайсироваться) в разных комбинациях. Более того, нуклеотидная последовательность, выступающая как экзон в одном случае, ведет себя как интрон в другом. Эти возможности выбора путей сплайсинга молекулы РНК-предшественника представлены на рис. 12.

Рис. 12. Схема альтернативного сплайсинга пре-мРНК.
Так, например, участок экзона 1а может вести себя как интрон. Другой случай демонстрирует образование двух разных РНК, несущих участки, кодирующие разные белковые структуры. Так, если экзон 2 кодирует в молекуле белка элементы α-спирали, а экзон 3 - так называемой β-структуры, то РНК 1 будет кодировать белок, не включающий указанный участок β-структуры, а РНК 2, напротив, служит матрицей при образовании белка без участка α-спирали. В результате образуются так называемые изоформы белка. Это наблюдаемое для многих генов явление выбора путей созревания мРНК получило название альтернативного сплайсинга. Многие гены содержат десяток и более экзонов. Поэтому число вариантов зрелых молекул мРНК, содержащих разные экзоны, потенциально может быть достаточно большим, приближаясь к величине 2n, где n - число экзонов. Таким образом, экзон-интронная структура оказывается чрезвычайно экономичной, обеспечивая большое разнообразие белков, кодируемых разными мРНК, которые, однако, произошли из одного и того же РНК-предшественника в результате транскрипции одного гена.
Случаи альтернативного сплайсинга имеют место в мышечной ткани, приводя к образованию функционально различающихся, но во многом сходных (содержат общие экзоны!) белков. Рассмотрим конкретный пример - тропомиозин, белок, участвующий в мышечном сокращении. В разных клетках с гена тропомиозина синтезируются разные белки:
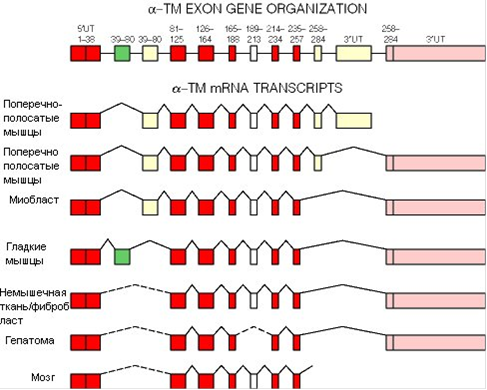
Это же явление широко распространено в нервной ткани, где, как известно, транскрибируется огромное множество генов, обеспечивая разнообразие белков. Так, например, разнообразие белков оболочки нервных волокон обусловлено альтернативным сплайсингом. Альтернативный сплайсинг нередко используется при синтезе вариантов белков - "факторов транскрипции", необходимых для включения (или выключения) гена и работы РНК-полимеразы. Наличие белка - фактора транскрипции может определять судьбу клетки, ее дифференцировку в клетку специализированной ткани: нервной, жировой, почечной и т.д. Альтернативный сплайсинг выполняет свою роль и при образовании вариантов белков клеточной адгезии, которые отвечают за передвижение клеток и структурирование тканей. Бактерии, обладающие, как правило, генами без интронов, лишены этого способа регуляции работы (экспрессии) генов.
Каким образом осуществляется выбор пути альтернативного сплайсинга, являющегося важным механизмом регуляции активности генов и клеточной дифференцировки? Характер сплайсинга регулируется белками, способными связываться с молекулой РНК в районах интронов или пограничных участках экзон-интрон. Такие белки могут блокировать удаление одних интронов, в то же время активируя вырезание других. РНК-предшественник складывается в сложную трехмерную структуру, взаимодействуя с другими компонентами сплайсосомы. Поэтому нетрудно представить себе такие пространственные изменения этой структуры при участии белков, регулирующих сплайсинг, которые исключат катализ объединения одной пары интронов и, напротив, будут эффективно способствовать сшиванию другой пары.
Направленная регуляция путей сплайсинга может иметь громадные биологические последствия, например определять пол организма. Случай определения пола, основанный на регуляции сплайсинга, достаточно хорошо изучен у плодовой мушки дрозофилы (рис.13). Успехи этих исследований основаны на работах генетиков, изучавших мутации, нарушающие становление пола. Огромную роль здесь сыграли также работы, в которых в экспериментах in vitro были использованы "клонированные гены" (рекомбинантные ДНК).
Перейти на страницу:
1 2 3