После связывания U2 мяРНП к сплайсосоме присоединяется крупный рибонуклеопротеид, содержащий 3 мяРНК: U4,U5 и U6 (рис. 6). В результате образуется комплекс B1, содержащий 5 молекул мяРНК. U1 мяРНК в этом комплексе связана с 5’SS, U2 - с местом разветвления, а U4 мяРНК связана с U6 многочисленными комплементарными взаимодействиями и тем самым до поры до времени блокирует активность U6 мяРНК. Все мяРНК, кроме U6 синтезируются РНК полимеразой II и имеют классические атрибуты мяРНП - связанные Sm белки и триметилированный кеп. U6 мяРНК транскрибируется РНК полимеразой III и не имеет кеп-структуры. Кроме того, вместо Sm белков с ней связаны гомологичные им Lsm белки.
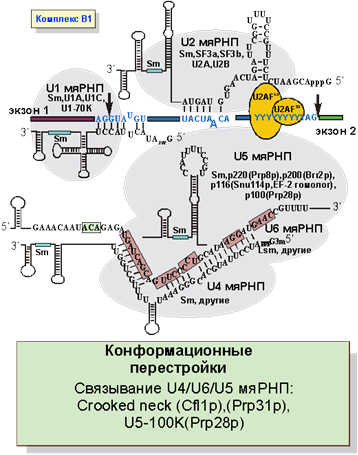
Рис. 6. Образование B1 комплекса: связывание U4/U5/U5 мяРНП.
После образования B1 комплекса происходит самая драматическая конформационная перестройка сплайсосомы (рис. 7). Во-первых, теряется связь U1 мяРНК с 5'SS. Во-вторых, расплетается дуплекс между U4 и U6 мяРНК. В-третьих, образуются дуплексы между U6 мяРНК и 5’SS, а также U6 и U2 мяРНК. Самую активную роль в этих перестройках играют белки, ассоциированные с U5 мяРНК. U5-200k расплетает дуплекс между U4 и U6 мяРНК, а U5-100k помогает образовать контакт между U6 и 5’SS. Другой белок, ассоциированный с U5 (U5-220k) способствует тому, что консервативная шпилька U5 мяРНК связывает 5’SS и 3’SS. В результате оказывается, что дуплекс U2 и U6 связывает место разветвления и 5'-место сплайсинга. U5 мяРНК дополнительно фиксирует 5’SS так, что оказывается возможным провести нуклеофильныю атаку 2’-OH выпетливающегося аденозина на межнуклеотидную связь 5’SS.
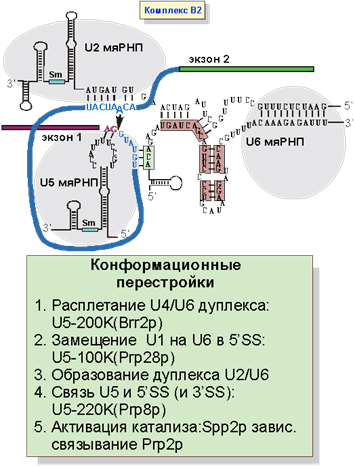
Рис. 7. Образование комплекса B2.
Переход от B2 комплекса к C1 как раз соответствует первой стадии сплайсинга, т.е. образованием лассообразной структуры и освобождению 3’-гидроксила 5’-концевого экзона (рис.8). В самом каталитическом акте сплайсосомы принимают участие белки Spp2p и Prp2p, хотя основную роль в катализе играют не белки, а сплайсосомная РНК.
После прохождения первой стадии сплайсинга необходимо подставить в активный центр сплайсосомы 3’SS. За выбор 3’-места сплайсинга отвечает U5-220k белок. Целый ряд белковых факторов играют роль в подстановке 3’-места сплайсинга в активный центр. Кроме того, осуществляется контроль за правильностью проведения первой стадии сплайсинга в котором принимает участие белок Prp16p.
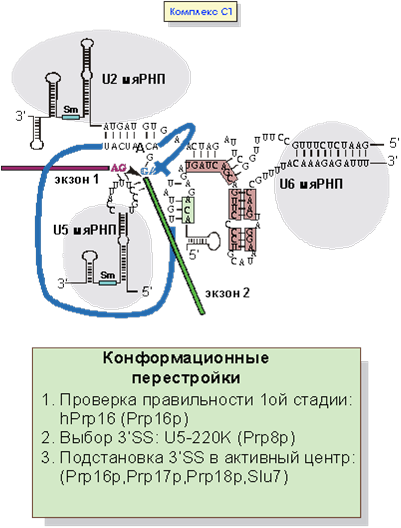
Рис. 8. Образование комплекса C1: подстановка 3’SS в активный центр.
Особую роль в позиционировании 5’-экзона и 3'-места сплайсинга играет U5 мяРНК. Молекула U5 мяРНК содержит петлю из неспаренных нуклеотидов на ножке (стволике), где нуклеотиды образуют пары, создавая двойную РНК-спираль. Нуклеотиды петли, взаимодействуя за счет водородных связей с экзонами, удерживают их вблизи друг от друга. Кроме того, первый и последний нуклеотиды интрона образуют неканонические взаимодействия двух G (гуанозинов). Эти взаимодействия помогают стягивать экзоны и осуществлять сплайсинг. Характер водородных связей функциональных групп гуанина отличается от классических, наблюдаемых в паре G-C Уотсона и Крика. Так, аминогруппа G, участвующая в установлении комплементарных взаимодействий с С, здесь остается вне игры. Вторая стадия сплайсинга, катализируемая сплайсосомой заключается в атаке 3’-гидроксила первого экзона на межнуклеотидную связь между интроном и 3’-концевым экзоном. В результате экзоны оказываются соединенными, а интрон - вырезанным в виде лассообразной структуры (рис.9).
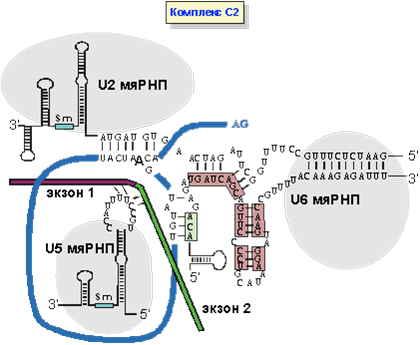
Рис. 9. Образование комплекса C2. Лигирование экзонов и полное выщепление интрона.
Перейти на страницу:
1 2 3