В 1977 г., к удивлению всего научного мира, да и к своему собственному, две группы исследователей - Ф. Шарп с коллегами в Массачусетском технологическом институте и группа сотрудников Колд-Спринг-Харборской лаборатории - открыли, что у высших организмов гены «разорваны». Оказалось, что кодирующие белок последовательности прерываются вставками, которые не кодируют белок и при созревании молекулы удаляются из предшественника РНК. Эти участки были названы интронами. Участки РНК, ковалентно соединяющиеся друг с другом и сохраняющиеся в составе зрелой РНК, называют экзонами. Зрелая информационная РНК, содержащая экзоны, транслируется в цитоплазме с участием рибосом. Оказалось, что экзон-интронная структура генов эукариот является скорее правилом, а гены без интронов - немногочисленными исключениями. Гены бактерий обычно не содержат экзонов. Экзоны включают последовательность нуклеотидов, содержащую следующие друг за другом кодоны (нуклеотидные триплеты), определяющие положение аминокислот в белке. Молекулы РНК, как и комплементарные нити двойной спирали ДНК, полярны, различают 5'- и 3'-концы молекулы. Участки экзонов или даже отдельные целые экзоны, располагающиеся на 5'- и 3'-концах молекулы информационной РНК, могут не использоваться при трансляции, они не кодируют аминокислоты.
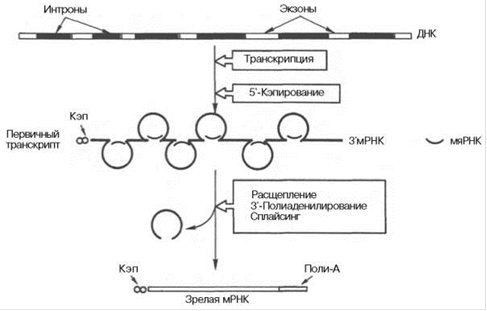
Рис.1. Биогенез мРНК у эукариот.
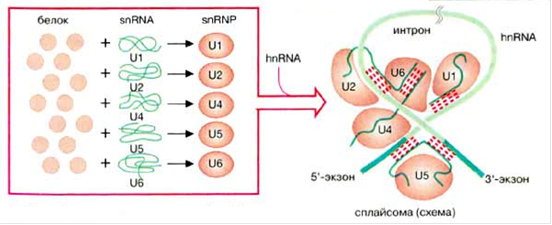
Рис.2. Схема мяРНП и сплайсосомы.
Как белки и РНК, осуществляющие сплайсинг, узнают какой фрагмент пре-мРНК нужно вырезать? И у дрожжей и у высших эукариот есть консервативные последовательности, определяющие границы интронов - GU на 5’- конце и AG на 3’- конце (рис. 3). Выделяют три функционально важных участка пре-мРНК, участвующие в сплайсинге: 5’-место сплайсинга (5’SS), 3’- место сплайсинга (3’SS) и место разветвления(BS), содержащее в
нуклеотидной последовательности A (аденин), который не образует пары с U и, следовательно, не входит в состав двойной спирали в результате взаимодействия с мяРНК. Поэтому этот А обладает особой реакционной способностью. На первой стадии 2’-гидроксил аденозина места разветвления атакует фосфодиэфирную связь 5’SS. В итоге этой атаки образуется структура “лассо” и свободный 3’-гидроксил 5’-концевого экзона. Во время второй стадии этот 3’-гидроксил атакует 3’SS. В результате экзоны оказываются ковалентно соединенными обычной межнуклеотидной связью, а интрон уходит в виде структуры “лассо”.
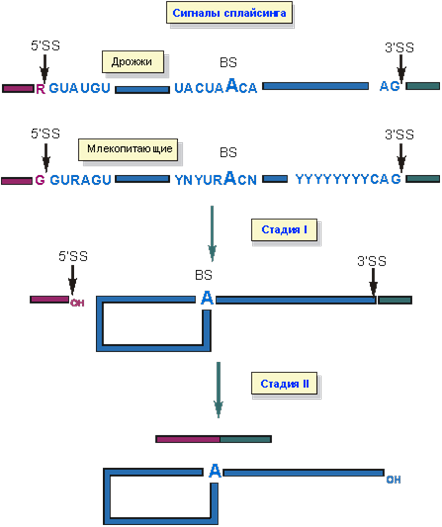
Рис. 3. Нуклеотидные последовательности, определяющие границы интронов. Схема изменения ковалентных связей при вырезании интрона.
Как же и в какой последовательности узнаются места сплайсинга и разветвления? Сначала U1 мяРНП узнает 5’-место сплайсинга, а возможно и 3’-место сплайсинга (рис. 4). Место разветвления узнается белком SF1 (на рисунках в скобках представлены названия соответствующих белков в дрожжах), а полипиримидиновая последовательность, предшествующая 3'SS - гетеродимером U2AF. При этом образуется комплекс CC (commitment complex) - комплекс определяущий вырезание данного интрона.
На следующем этапе комплекс CC переходит в комплекс A (рис. 5). Белок SF1 вытесняется с помощью UAP56 и с местом разветвления связывается U2 мяРНП. В самом U2 мяРНП фрагмент U2 мяРНК, комплементарный BS, освобождается от белка SAP61. Связыванию U2 РНП помогает белок U2AF.
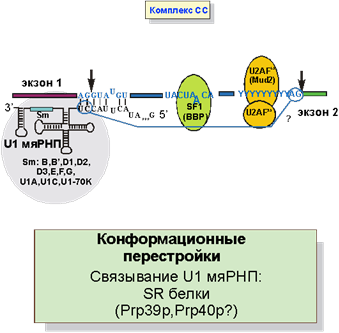
Рис. 4. Первичное узнавание интрона: образование комплекса CC.
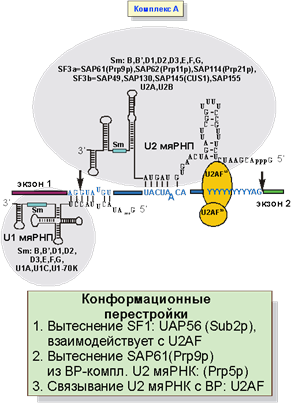
Рис. 5. Образование комплекса A: присоединение U2 мяРНП.
Перейти на страницу:
1 2 3