В состав молекулы гемоглобина человека входят две a-цепи (a-цепь закодирована в 16-ой хромосоме) и две b-цепи (b-цепь закодирована в 11-ой хромосоме). В состав b-цепи входит 146 аминокислотных остатков, при этом в нормальной b-цепи шестым аминокислотным остатком является глутаминовая кислота. С участием нормальной b-цепи образуется нормальный гемоглобин - HbA. В нетранскрибируемой нити участка ДНК, кодирующего b-цепь, глутаминовая кислота закодирована триплетом ГАА. Если же в результате мутации в ДНК произойдет замена триплета ГАА на триплет ГТА, то на месте глутаминовой кислоты в молекуле гемоглобина в соответствии с генетическим кодом появится валин. В итоге вместо гемоглобина HbA появится новый гемоглобин - HbS. Такая замена всего лишь одного нуклеотида и одной аминокислоты приводит к развитию тяжелого заболевания - серповидноклеточной анемии.
Различают несколько типов аномальных гемоглобинов.
. Изменена наружная часть молекулы. Почти все замены аминокислот на поверхности молекулы гемоглобина безвредны. Гемоглобин S представляет собой поразительное исключение.
. Изменен активный центр. В этом случае в дефектной субъединице не происходит связывания кислорода, так как структурные изменения вблизи гемма непосредственно влияют на связывание кислорода.
. Изменена третичная структура. Замена аминокислот препятствует возникновению нормальной конформации молекулы. Такие гемоглобины обычно нестабильны.
. Изменена четвертичная структура. Некоторые мутации, затрагивающие участки области контактов, приводят к потере аллостерических свойств. В результате нарушается сродство таких гемоглобинов к О2.
В результате некоторых мутаций гемоглобины утрачивают стабильность из-за деформаций третичной структуры
Замены аминокислот в отдаленных от гемма участках могут препятствовать возникновению нормальной конформации гемоглобина и тем самым резко нарушать его функцию. Большой интерес в этом отношении представляет гемоглобин Riverdale-Bronx, у которого в положении В6 вместо глицина стоит аргинин. Из-за больших размеров аргинин не умещается в том узком пространстве, в котором должны пересекаться спирали В и Е (рис. 3). Действительно, во всех нормальных гемоглобинах и миоглобинах положение В6 всегда занято глицином. Гемоглобин Riverdale-Bronx в результате произошедшей аминокислотной заметы оказывается нестабильным.
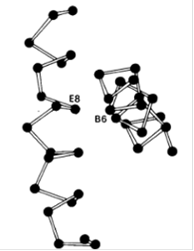
Рис. 3. Молекула гемоглобина принимает нормальную структуру, только если в положении В6 стоит остаток глицина. Поскольку именно в этом участке перекрещиваются спирали В и Е, для большей, чем у глицина, боковой цепи нет места.
Клиническая картина талассемии может возникать при аминокислотных заменах с образованием нестабильных гемоглобинов, при мутациях в терминирующем кодоне, приводящих к синтезу аномально длинных цепей гемоглобина (например, при гемоглобинопатий Constant Spring ) или при слиянии генов дельта- и бета-цепей, приводящих к синтезу аномально коротких цепей гемоглобина (например, при гемоглобинопатий Lepore ). При гемоглобинопатий E точечная мутация не только приводит к замене аминокислоты, но и нарушает сплайсинг транскрипта.
Цепи глобина принято обозначать греческими буквами, а мутации - буквами в верхнем регистре. Например, A в верхнем регистре используется для обозначения нормального гемоглобина, а относительно широко распространенный гемоглобин S нередко обозначается как бета с буквой s в верхнем регистре. Чаще, однако, указывается, какая замена аминокислоты произошла, и тогда гемоглобин S обозначается как бета(6Глу-Вал), при этом, естественно (6Глу-Вал) находится в верхнем регистре.
Задание № 7
Репликация ДНК-механизм, основные этапы. Ферменты и механизм репарации ДНК. Ингибиторы репликации как противоопухолевые и противовирусные препараты, их механизмы действия (привести не менее 5 примеров). Ответить на вопрос: какой результат получили бы Мезельсон и Сталь, если бы репликация была бы консервативной. Укажите предполагаемое распределение молекул ДНК после 1-й и 2-й генераций в случае консервативной репликации (т.е. когда родительская двойная спираль не расходится)?
Реплика́ция ДНК - это процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты, который происходит в процессе деления клетки на матрице родительской молекулы ДНК. При этом генетический материал, зашифрованный в ДНК, удваивается и делится между дочерними клетками. Репликацию ДНК осуществляет фермент ДНК-полимераза.
Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у эукариот осуществляется несколькими разными ДНК-полимеразами. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.