В зависимости от длины саркомеров длина участков, в которых нити актина и миозина перекрываются друг с другом, будет различной и, следовательно, будет различно количество мостиков, участвующих и создании напряжения, развиваемого мышцей. Учитывая, что максимальная сила миофибриллы определяется количеством функционирующих мостиков, следует ожидать, что максимальная сила изометрического сокращения миофибриллы будет изменяться с изменением длины саркомера. При длине саркомера 3,65 мкм нити актина и миозина уже не накладываются друг на друга и можно ожидать, что волокно не будет способно развивать силу. Под силой сокращения следует понимать разность между общей силой, развиваемой при раздражении мышцей, и упругой восстанавливающей силой, обусловленной эластическими элементами мышцы в случае се растяжения сверх нормальной длины. По мере сближения Z-мембран нити актина все глубже проникают в промежутки между нитями миозина и, наконец, при расстоянии 2,2 мкм все мостики миозиновых нитей приходят в контакт с нитью актина. Если именно эти мостики ответственны за возникновение силы, то следует ожидать, что в диапазоне от положения I до положения II сила будет пропорциональна степени перекрывания нитей. При дальнейшем укорочении волокна число мостиков, которые могут замыкаться, не изменяется и сила должна оставаться постоянной, пока длина саркомера не уменьшится до 2,05 мкм. В этот момент нити актина сходятся своими концами и сила должна убывать вследствие того, что тонкие нити, которые проникли дальше середины А-диска, будут неправильно ориентированы по отношению к миозиновым мостикам. Сила должна постепенно убывать, пока расстояние не достигнет 1,65 мкм, когда концы миозиновых нитей приходят в соприкосновение с Z-мембранами. При дальнейшем сокращении нити миозина должны деформироваться; сила должна убывать быстрее и совсем исчезать, когда актиновые нити доходят до противолежащих Z-мембран.
Все эти предположения подтвердились экспериментально. Гордоном, А.Хаксли, Юлианом (1966) измерялось напряжение, развиваемое мышечным волокном при изометрическом сокращении, и одновременно методом фазово-контрастной микроскопии регистрировалась длина саркомера.
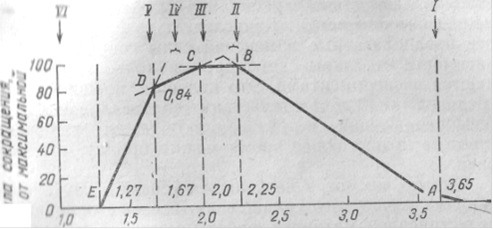
Рис.4. Зависимость силы сокращения поперечнополосатого мышечного волокна от длины саркомера
Однако, несмотря на большие успехи в изучении механизма мышечного сокращения, все еще окончательно не установлен механизм работы мостиков, в результате которой энергия гидролиза АТФ превращается в механическую работу.
В настоящее время имеется ряд гипотез, пытающихся объяснить конкретный механизм взаимодействия актиновых и миозиновых нитей.
Наиболее глубоко разработанной и обоснованной является гипотеза Дэвиса. Согласно этой гипотезе, мостик между миозиновой и актиновой нитями образован полипептидными цепочками конца миозиновой молекулы, скрученными в спираль. В покое мостик вытянут-спираль находится в растянутом состоянии. Это обусловлена электростатическим отталкиванием двух отрицательных зарядов. Один из них находится в фиксированном состоянии у основания мостика, которое обладает АТФ-азной активностью. Другой отрицательный заряд локализован па конце мостика, с которым связана молекула АТФ.
При возбуждении мышцы саркоплазматический ретикулум освобождает ионы кальция. Они образуют связь между молекулой АТФ, находящейся на конце мостика, и молекулой АДФ, расположенной на актиновой нити, что вызывает нейтрализацию отрицательных зарядов. Электростатическое отталкивание исчезает и растянутая цепочка - мостик - скручивается в α-спираль благодаря образованию водородных связей. Этот процесс представляет собой освобождение потенциальной энергии, запасенной вытянутой полипептидной цепочкой при первоначальном отталкивании зарядов. Укорочение полипептидной цени с образованием α-спирали приводит к двум эффектам. Во-первых, актиновая нить перемещается относительно миозиновой на один шаг; во-вторых, присоединенная молекула АТФ перемещается в область гипотетического АТФ-азного центра. Благодаря соответствующему расположению этого центра и наклону мостиков относительно толстой нити актиновые нити перемещаются в сторону М-линий. После этого АТФ расщепляется на АДФ и минеральный фосфат, что ведет к разрыву связей между актином и миозином. На место молекулы АДФ в миозиновом мостике из саркоплазмы поступает новая молекула АТФ, которая отталкивается отрицательным фиксированным зарядом миозина. В результате этого α-спираль растягивается - мостик удлиняется. Если в саркоплазме в это время имеются свободны ионы кальция, то весь цикл повторяется сначала.
Перейти на страницу:
1 2 3 4 5 6 7