Мышечные сокращения делятся на изометрические
- происходящие при неизменной длине мышцы, и изотонические
- происходящие при неизменном напряжении. Чисто изометрические или чисто изотонические сокращения с большим или меньшим приближением можно получить только в лабораторных условиях при работе на изолированных мышцах. В организме сокращения мышц никогда не бывают чисто изометрическими или чисто изотоническими.
Нарколог на дом вывод из запоя Аб Клиника
Скелетные мышцы с помощью сухожилий прикрепляются к костям, которые образуют систему рычагов. В большинстве случаев мышцы прикрепляются к костям так, что при их сокращении наблюдается выигрыш в амплитуде движений и эквивалентный проигрыш в силе. Плечо рычага мышцы в большинстве случаев бывает меньше плеча рычага соответствующей кости. Согласно Аккерману, механический выигрыш в амплитуде движений большинства конечностей человека имеет величину от 2,5 до 20. Для двуглавой мышцы плеча он равен приблизительно 10. При движении костей соотношение плеч рычагов мышц и костей меняется, что приводит к изменению напряжения мышц. По этой причине изотонических сокращений в естественных условиях не наблюдается. По этой же причине в процессе сокращения меняются вышеприведенные величины механического выигрыша в амплитуде движений.
В зависимости от величины силы, которую преодолевает мышца, скорость сокращения (укорочения) мышцы бывает различной. Хилл на основе опытных данных, полученных при работе на изолированных мышцах, вывел так называемое основное уравнение сокращения мышцы. Согласно Хиллу, скорость сокращения мышцы v находится в гиперболической зависимости от величины нагрузки F:
(F + a) (v + b) = const,
где а и b - константы, приблизительно равные ¼ F и соответственно ¼ v.
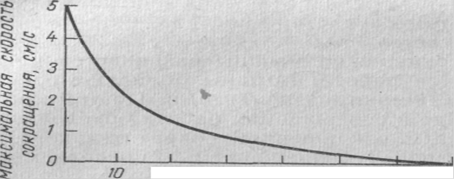
Нагрузка, кгс
Рис.1. Зависимость скорости сокращения мышцы лягушки от величины нагрузки
Байером были сделаны интересные замечания к уравнению. Уравнение приводится к виду
F' v' = const,
если принять F' = F + a и v' = v + b. Произведение F х v' представляет собой общую мощность, развиваемую мышцей при сокращении. Так как Fv меньше F'v', т. е. внешняя мощность меньше общей мощности, то следует предположить, что мышца совершает не только внешнюю работу, но еще и некоторую внутреннюю работу, проявляющуюся в том, что нагрузка как бы увеличивается на величину а, а скорость сокращения на величину b. Эту внутреннюю работу можно интерпретировать как потерю энергии на внутримолекулярное трение в форме теплового рассеивания. Тогда с учетом высказанных замечаний можно отметить, что общая мощность мышцы в физиологических пределах является постоянной величиной, не зависящей от величины нагрузки и скорости сокращения.
С термодинамической точки зрения мышца представляет собой систему, которая преобразует химическую энергию (энергию АТФ) в механическую работу, т. е. мышца является хемо-механической машиной.
Как уже отмечалось, при сокращении мышцы происходит теплообразование. Хиллом с помощью термоэлектрических методов было установлено, что при каждом раздражении вначале выделяется постоянная по величине и не зависящая от нагрузки теплота активации Q, а затем теплота сокращения kΔl, пропорциональная сокращению мышцы Δl и не зависящая от нагрузки (k-коэффициент пропорциональности). Если сокращение изотоническое, то мышца производит работу А, равную произведению нагрузки F на величину сокращения: А = FΔl. Согласно первому закону термодинамики, изменение внутренней энергии ΔU мышцы будет равняться сумме выделенного тепла и совершенной работы:
Перейти на страницу:
1 2 3