Рибосомные РНК синтезируются в виде большого предшественника. Этот предшественник нарезается на зрелые молекулы рРНК с помощью эндо- и экзонуклеаз. Кроме того, отдельные нуклеотиды рРНК модифицируются.
Предшественник рРНК прокариот содержит фрагменты, соответствующие 16S, 23S и 5S рРНК. Как правило, в этом транскрипте содержатся также несколько пре-тРНК. Большие рибосомальные РНК образуют домены подобного транскрипта. Их 5’ и 3’-концы сближены за счет комплементарности прилегающих участков РНК. Такая двухцепочечная структура разрезается РНКазой III, а оставшиеся несколько лишних нуклеотидов отрезаются экзонуклеазами. Модификация оснований рРНК проводится набором ферментов, причем один фермент может модифицировать от 1 до 3 нуклеотидных остатков.
У эукариот созревание также включает в себя разрезание предшественника (рис. 14), причем кроме 18S, 28S, и 5S рРНК появляется еще и 5.8S рРНК. Кроме различных эндо- и экзонуклеаз, в созревании принимают участие малые ядрышковые РНК (мяРНК). Эти РНК локализованы преимущественно в ядрышке - участке ядра, где происходит транскрипция генов рРНК и сборка пре-рибосом.
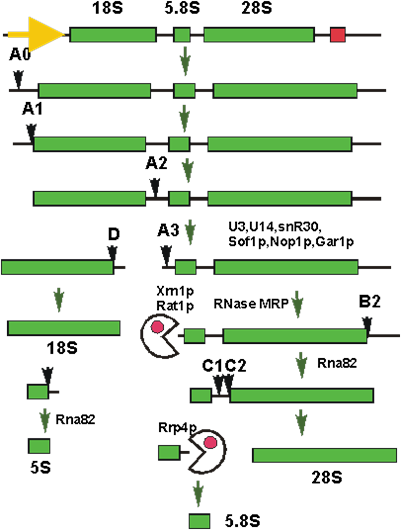
Рис.14. Схема процесса разрезания предшественника рРНК эукариот.
Если нарезание предшественника рРНК, в общем, сходно у про- и эукариот, то система модификации оснований рРНК у эукариот уникальна. В отличии от прокариот, у высших организмов модифицировано гораздо больше нуклеотидных остатков рРНК. Среди модификаций подавляющее большинство это псевдоуридилирование и 2’-O-метилирование рибозы. Невозможно представить, чтобы модификации всех этих остатков осуществлялись каждая своим ферментом.
Оказалось, что в ядрышке, помимо мяРНК, участвующих в разрезании пре-рРНК, существует множество других мяРНК. Эти небольшие РНК можно разделить на 2 класса: C/D и H/ACA (рис. 15).

Рис. 15. Свойства мяРНК различных классов.
МяРНК обоих классов транскрибируются РНК пролимеразой II, но не как самостоятельные гены, а в интронах других генов. Часто мяРНК входят в состав интронов мРНК рибосомных белков. Есть случаи, когда единственной функцией некоторых генов служит производство мяРНК, содержащихся в интронах. При этом открытая рамка считывания зрелой мРНК прервана многочисленными стоп-кодонами и не кодирует какого-либо белкового продукта.
Характерная особенность мяРНК - короткие участки комплементарности различным областям рРНК. У РНК C/D класса эта область комплементарности более 10 нуклеотидов, а у H/ACA - меньше. С каждым классом мяРНК связан свой набор белков. мяоРНП C/D класса осуществляют 2’-O-метилирование, а H/ACA - псевдоуридилирование рРНК.
Структура мяРНК представлена на рисунке 17. 5’- и 3’-концы C/D мяРНК спарены. Следом расположены две консервативные последовательности нуклеотидов, из-за которых этот класс и получил свое название. Сразу за D последовательностью идет участок комплементарности рРНК. Третий нуклеотид рРНК от участка начала комплементарности 2’-O-метилируется. У H/ACA мяРНК также имеется две консервативные последовательности, давшие название этому классу. Нуклеотиды, комплементарные рРНК располагаются в двух внутренних шпильках. Остаток уридина в положении, отмеченном на рисунке 16, изомеризуется в псевдоуридин.
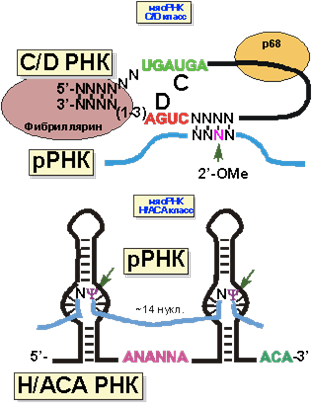
Рис. 16. Модель структуры комплексов мяРНК-рРНК, в которых происходит модификация нуклеотидов.
С помощью мяРНК эукариотические организмы решают проблему специфичной модификации остатков рРНК, без привлечения огромного числа специфичных ферментов. Кстати, малые РНК, подобные мяРНК участвуют в модификации малых ядерных РНК.